Follow infophilosopher:
 |
|

|
|
|
Philosophers
Mortimer Adler Rogers Albritton Alexander of Aphrodisias Samuel Alexander William Alston Anaximander G.E.M.Anscombe Anselm Louise Antony Thomas Aquinas Aristotle David Armstrong Harald Atmanspacher Robert Audi Augustine J.L.Austin A.J.Ayer Alexander Bain Mark Balaguer Jeffrey Barrett William Barrett William Belsham Henri Bergson George Berkeley Isaiah Berlin Richard J. Bernstein Bernard Berofsky Robert Bishop Max Black Susanne Bobzien Emil du Bois-Reymond Hilary Bok Laurence BonJour George Boole Émile Boutroux Daniel Boyd F.H.Bradley C.D.Broad Michael Burke Jeremy Butterfield Lawrence Cahoone C.A.Campbell Joseph Keim Campbell Rudolf Carnap Carneades Nancy Cartwright Gregg Caruso Ernst Cassirer David Chalmers Roderick Chisholm Chrysippus Cicero Tom Clark Randolph Clarke Samuel Clarke Anthony Collins Antonella Corradini Diodorus Cronus Jonathan Dancy Donald Davidson Mario De Caro Democritus Daniel Dennett Jacques Derrida René Descartes Richard Double Fred Dretske John Earman Laura Waddell Ekstrom Epictetus Epicurus Austin Farrer Herbert Feigl Arthur Fine John Martin Fischer Frederic Fitch Owen Flanagan Luciano Floridi Philippa Foot Alfred Fouilleé Harry Frankfurt Richard L. Franklin Bas van Fraassen Michael Frede Gottlob Frege Peter Geach Edmund Gettier Carl Ginet Alvin Goldman Gorgias Nicholas St. John Green H.Paul Grice Ian Hacking Ishtiyaque Haji Stuart Hampshire W.F.R.Hardie Sam Harris William Hasker R.M.Hare Georg W.F. Hegel Martin Heidegger Heraclitus R.E.Hobart Thomas Hobbes David Hodgson Shadsworth Hodgson Baron d'Holbach Ted Honderich Pamela Huby David Hume Ferenc Huoranszki Frank Jackson William James Lord Kames Robert Kane Immanuel Kant Tomis Kapitan Walter Kaufmann Jaegwon Kim William King Hilary Kornblith Christine Korsgaard Saul Kripke Thomas Kuhn Andrea Lavazza James Ladyman Christoph Lehner Keith Lehrer Gottfried Leibniz Jules Lequyer Leucippus Michael Levin Joseph Levine George Henry Lewes C.I.Lewis David Lewis Peter Lipton C. Lloyd Morgan John Locke Michael Lockwood Arthur O. Lovejoy E. Jonathan Lowe John R. Lucas Lucretius Alasdair MacIntyre Ruth Barcan Marcus Tim Maudlin James Martineau Nicholas Maxwell Storrs McCall Hugh McCann Colin McGinn Michael McKenna Brian McLaughlin John McTaggart Paul E. Meehl Uwe Meixner Alfred Mele Trenton Merricks John Stuart Mill Dickinson Miller G.E.Moore Ernest Nagel Thomas Nagel Otto Neurath Friedrich Nietzsche John Norton P.H.Nowell-Smith Robert Nozick William of Ockham Timothy O'Connor Parmenides David F. Pears Charles Sanders Peirce Derk Pereboom Steven Pinker U.T.Place Plato Karl Popper Porphyry Huw Price H.A.Prichard Protagoras Hilary Putnam Willard van Orman Quine Frank Ramsey Ayn Rand Michael Rea Thomas Reid Charles Renouvier Nicholas Rescher C.W.Rietdijk Richard Rorty Josiah Royce Bertrand Russell Paul Russell Gilbert Ryle Jean-Paul Sartre Kenneth Sayre T.M.Scanlon Moritz Schlick John Duns Scotus Arthur Schopenhauer John Searle Wilfrid Sellars David Shiang Alan Sidelle Ted Sider Henry Sidgwick Walter Sinnott-Armstrong Peter Slezak J.J.C.Smart Saul Smilansky Michael Smith Baruch Spinoza L. Susan Stebbing Isabelle Stengers George F. Stout Galen Strawson Peter Strawson Eleonore Stump Francisco Suárez Richard Taylor Kevin Timpe Mark Twain Peter Unger Peter van Inwagen Manuel Vargas John Venn Kadri Vihvelin Voltaire G.H. von Wright David Foster Wallace R. Jay Wallace W.G.Ward Ted Warfield Roy Weatherford C.F. von Weizsäcker William Whewell Alfred North Whitehead David Widerker David Wiggins Bernard Williams Timothy Williamson Ludwig Wittgenstein Susan Wolf Xenophon Scientists David Albert Michael Arbib Walter Baade Bernard Baars Jeffrey Bada Leslie Ballentine Marcello Barbieri Gregory Bateson Horace Barlow John S. Bell Mara Beller Charles Bennett Ludwig von Bertalanffy Susan Blackmore Margaret Boden David Bohm Niels Bohr Ludwig Boltzmann Emile Borel Max Born Satyendra Nath Bose Walther Bothe Jean Bricmont Hans Briegel Leon Brillouin Stephen Brush Henry Thomas Buckle S. H. Burbury Melvin Calvin Donald Campbell Sadi Carnot Anthony Cashmore Eric Chaisson Gregory Chaitin Jean-Pierre Changeux Rudolf Clausius Arthur Holly Compton John Conway Simon Conway-Morris Jerry Coyne John Cramer Francis Crick E. P. Culverwell Antonio Damasio Olivier Darrigol Charles Darwin Richard Dawkins Terrence Deacon Lüder Deecke Richard Dedekind Louis de Broglie Stanislas Dehaene Max Delbrück Abraham de Moivre Bernard d'Espagnat Paul Dirac Hans Driesch John Dupré John Eccles Arthur Stanley Eddington Gerald Edelman Paul Ehrenfest Manfred Eigen Albert Einstein George F. R. Ellis Hugh Everett, III Franz Exner Richard Feynman R. A. Fisher David Foster Joseph Fourier Philipp Frank Steven Frautschi Edward Fredkin Augustin-Jean Fresnel Benjamin Gal-Or Howard Gardner Lila Gatlin Michael Gazzaniga Nicholas Georgescu-Roegen GianCarlo Ghirardi J. Willard Gibbs James J. Gibson Nicolas Gisin Paul Glimcher Thomas Gold A. O. Gomes Brian Goodwin Joshua Greene Dirk ter Haar Jacques Hadamard Mark Hadley Patrick Haggard J. B. S. Haldane Stuart Hameroff Augustin Hamon Sam Harris Ralph Hartley Hyman Hartman Jeff Hawkins John-Dylan Haynes Donald Hebb Martin Heisenberg Werner Heisenberg Grete Hermann John Herschel Basil Hiley Art Hobson Jesper Hoffmeyer Don Howard John H. Jackson William Stanley Jevons Roman Jakobson E. T. Jaynes Pascual Jordan Eric Kandel Ruth E. Kastner Stuart Kauffman Martin J. Klein William R. Klemm Christof Koch Simon Kochen Hans Kornhuber Stephen Kosslyn Daniel Koshland Ladislav Kovàč Leopold Kronecker Rolf Landauer Alfred Landé Pierre-Simon Laplace Karl Lashley David Layzer Joseph LeDoux Gerald Lettvin Gilbert Lewis Benjamin Libet David Lindley Seth Lloyd Werner Loewenstein Hendrik Lorentz Josef Loschmidt Alfred Lotka Ernst Mach Donald MacKay Henry Margenau Owen Maroney David Marr Humberto Maturana James Clerk Maxwell Ernst Mayr John McCarthy Warren McCulloch N. David Mermin George Miller Stanley Miller Ulrich Mohrhoff Jacques Monod Vernon Mountcastle Emmy Noether Donald Norman Travis Norsen Alexander Oparin Abraham Pais Howard Pattee Wolfgang Pauli Massimo Pauri Wilder Penfield Roger Penrose Steven Pinker Colin Pittendrigh Walter Pitts Max Planck Susan Pockett Henri Poincaré Daniel Pollen Ilya Prigogine Hans Primas Zenon Pylyshyn Henry Quastler Adolphe Quételet Pasco Rakic Nicolas Rashevsky Lord Rayleigh Frederick Reif Jürgen Renn Giacomo Rizzolati A.A. Roback Emil Roduner Juan Roederer Jerome Rothstein David Ruelle David Rumelhart Robert Sapolsky Tilman Sauer Ferdinand de Saussure Jürgen Schmidhuber Erwin Schrödinger Aaron Schurger Sebastian Seung Thomas Sebeok Franco Selleri Claude Shannon Charles Sherrington Abner Shimony Herbert Simon Dean Keith Simonton Edmund Sinnott B. F. Skinner Lee Smolin Ray Solomonoff Roger Sperry John Stachel Henry Stapp Tom Stonier Antoine Suarez Leo Szilard Max Tegmark Teilhard de Chardin Libb Thims William Thomson (Kelvin) Richard Tolman Giulio Tononi Peter Tse Alan Turing C. S. Unnikrishnan Nico van Kampen Francisco Varela Vlatko Vedral Vladimir Vernadsky Mikhail Volkenstein Heinz von Foerster Richard von Mises John von Neumann Jakob von Uexküll C. H. Waddington James D. Watson John B. Watson Daniel Wegner Steven Weinberg Paul A. Weiss Herman Weyl John Wheeler Jeffrey Wicken Wilhelm Wien Norbert Wiener Eugene Wigner E. O. Wilson Günther Witzany Stephen Wolfram H. Dieter Zeh Semir Zeki Ernst Zermelo Wojciech Zurek Konrad Zuse Fritz Zwicky Presentations Biosemiotics Free Will Mental Causation James Symposium |
Aaron Schurger
Aaron Schurger earned his neuroscience Ph.D. from Princeton, working in Jonathan Cohen's Neuroscience of Cognitive Control Laboratory (NCC Lab). He is currently in France at the Cognitive Neuroimaging Unit of the Institut National de la Santé et de la Recherche Médicale (INSERM).
Cohen and Joshua Greene (a former NCC Lab researcher and now professor of psychology at Harvard University) published an important paper challenging conscious will, with the evidence based primarily on Benjamin Libet's famous experiments.
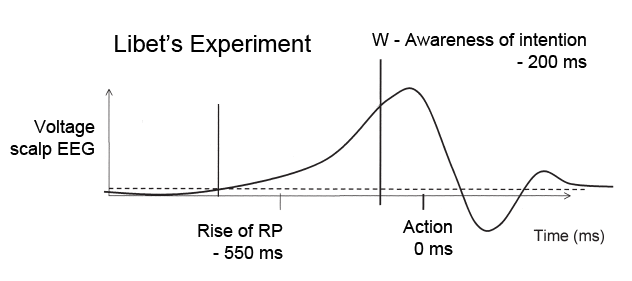
Now Schurger and his French colleagues, Jacobo D. Sitt and Stanislas Dehaene, have produced significant evidence that the readiness potential (RP) seen before voluntary self-initiated movements is not necessarily causal and determinative of the action.
In their 2012 article in the Proceedings of the National Academy of Sciences, they say:
The premovement buildup of neuronal activity apparent in the RP and the assumption of causality invested in it have become a cornerstone in the study of volition. Notably, Benjamin Libet (9, 15) tried to measure the temporal relationship between the onset of the RP and the feeling of an “urge” to move. The results of Libet et al.’s experiments (9, 15) suggested that the objective neural events in the brain that cause movement precede the urge to move by 300 ms or more. A recent experiment, using Libet’s paradigm, confirms the same preurge buildup at the single-neuron level (16). Such demonstrations have had an unrivaled influence on the prevailing view that movement is initiated preconsciously and the feeling of intending to move is grafted on after the fact. In fact a gradual increase in neural activity preceding spontaneous movements appears to be a very general phenomenon, common to both vertebrates (8, 16, 17) and invertebrates (18) alike. Why do both humans and crayfish (18) exhibit the same 1- to 2-s buildup of neural activity in advance of self-initiated movements? Kornhuber and Deecke’s (12) interpretation of the RP as a sign of planning and preparation for movement fails to explain what specific neural operations underlie the spontaneous self-initiation of movement and why these operations are reflected in the specific exponential shape of the RP.Schurger et al. find that the shape of the readiness potential can be explained if the brain uses a common machinery for decision making, specifically a threshold applied to the output of a stochastic neural accumulator. they say: Decision-making tasks are typically modeled in terms of the accumulation of evidence. What is unique to the spontaneous-movement task is that subjects are specifically instructed to not base their movement decisions on any specific evidence, sensory or otherwise. One simple solution, given these instructions, is to apply the same accumulator-plus-threshold decision mechanism, but fed solely with internal physiological noise.The stochastic-decision model reproduces the distribution of waiting times as well as the characteristic shape and time course of the readiness potential. Schurger et al. repeated the Libet experiments, but also added a variation that they call Libetus Interruptus. In the classic Libet experiment, the subject observes a rotating clock dial and notices the clock's position when the subject, without preplanning, flicks a wrist at a random time. The Libetus interruptus task is identical to the classic Libet task except for the addition of random interruptions: an audible “click” that cues the subject to make the movement as quickly as possible after the click. Recall that philosopher Alfred Mele is a powerful critic of interpretations of the Libet experiments which claim our decisions are made in our brain long before our conscious awareness of the decisions. Mele criticizes the interpretation of the Libet results on two grounds. First, the mere appearance of the RP a half-second or more before the action in no way makes the RP the cause of the action. It may simply mark the beginning of forming an intention to act. In the two-stage model, the rise of the RP might simply reflect the considering of possible options. Secondly, Libet himself argued that there is enough time after the W moment (a window of opportunity) to veto the action, but Mele's second criticism points out that such examples of "free won't" would not be captured in the classic Libet experiments, because the recording device is triggered by the action (typically flicking the wrist) itself. Thus, although all Libet experiments ended with the wrist flicking, we are not justified in assuming that the rise of the RP (well before the moment of conscious will) is a cause of the wrist flicking. Libet knew that there were very likely other times when the RP rose, but which did not lead to a flick of the wrist, so his experiment could not detect them. Schurger et al. note this backward selection bias, that only epochs ending with an actual movement are subject to analysis (ibid, p.6). Their Libetus interruptus is exploring those time intervals when the RP might rise, their accumulator model might get to, or even surpass?, the threshold, and yet there might be no wrist flick. They offer a new model for what the RP represents, beyond the vague phrases of the past four decades of research, that it reflects "planning and preparation for movement." Their model for the RP is divided into two nonlinear components: an early precommitment phase (or stage in our terminology) dominated by stochastic fluctuations (with an evolving spatial distribution) and a late postcommitment motor execution phase (the last 150 ms). Schurger et al. challenge the notion that the early buildup of activity biases supposedly "voluntary" decisions (as argued by Soon et al., among others) They say that their model is consistent with such predecision biases, but suggests that they may reflect stochastic fluctuations rather than an intentional (preconscious) decision process: It is widely assumed that the neural decision to move coincides with the onset of the RP (which, given its slow nonlinear character, is difficult to pinpoint) (11). Our model challenges that assumption by suggesting that the “neural decision to move now” might come very late in the time course of the RP. Prior research shows an involvement of motor areas, including primary motor cortex, in motor imagery, in the absence of overt movement (40). Thus, movement-specific activity in motor cortex, even primary motor cortex, although it might vary with the probability that a movement will occur, does not necessarily signal the final commitment to produce a movement now...Thus, according to our model, uncued movements in a task like Libet’s tend to be preceded by a gradual increase in neural activity [measured at the scalp (8, 9) or the single-neuron level (16)] whose causal role is incidental—not directed (consciously or otherwise) at producing a movement."Finally," say Schurger et al., although their "model is silent with respect to the urge to move and its temporal relation to motor decisions, it helps dissolve another puzzling question that seemed to arise from Libet’s paradigm. Libet himself found that subjects were able to estimate the time of a tactile sensory decision in relation to a quickly rotating clock dial with only about 50 ms of error on average (9). Why then should there be such a long and variable gap between the time of a motor decision and the subjective estimate of the time of the motor decision, whereas no such gap exists for sensory decisions? In fact, this question arises only when we assume that the motor decision coincides in time with the onset of the RP. We have argued that this need not be the case and that the neural decision to move may come much closer in time to the movement itself (e.g., −150 ms). We propose that the neural decision to move coincides in time with average subjective estimates of the time of awareness of intention to move (9, 11) and that the brain produces a reasonably accurate estimate of the time of its movement-causing decision events." (ibid, p.7) Thinkers (e.g., Daniel Wegner, Patrick Haggard) who claim that the Libet experiments prove that our conscious will and subsequent actions are caused by prior neural activity - this is the popular view that "my neurons made me do it" - are simply wrong. Note that the abrupt and rapid decisions to flex a finger measured by Libet bear little resemblance to the kinds of two-stage deliberate decisions for which we can first freely generate alternative possibilities for action, then evaluate and select (in an adequately causal way) which is the best of these possibilities in the light of our reasons, motives, and desires - first "free," then "will." 
For Teachers
For Scholars
|